 Embryologi utfordringer
Embryologi utfordringer
(fra 'Theistic Evolution - a Scientific, Philosophical and Theological critique, kap.8)
 "Ett av de vedvarende mysterier i utviklings- og celle -biologi, er morfogenese, prosessen av hvordan den arvede kroppsformen blir dannet på hvert stadium, fra det befruktede egget" (R. Keller)
"Ett av de vedvarende mysterier i utviklings- og celle -biologi, er morfogenese, prosessen av hvordan den arvede kroppsformen blir dannet på hvert stadium, fra det befruktede egget" (R. Keller)
"Problemet med biologisk form forblir uløst, til tross for de kjente detaljer med gen-aktivitet i embryo-utvikling." (Brian Goodwin)
"Å dechiffrere og lære å kontrollere form er ... uten tvil det fundamentale problemet i biologi og medisin." (Michael Levin; 7)
Men om mekanismene, gjennom hvilke, utviklingsprogram oversetter til form gjenstår å bli oppdaget, så forblir mekanismene som underligger stor-skala evolusjonære endringer høyst spekulative, siden disse essensielt avhenger av fundamentale endringer i den 'svarte boksen' til utviklingsprogrammet.
Om vi flytter søkelyset fra møte med det nye til 'embryologi' scenen', ser vi molekylære mønstre møte hverandre tilfeldig eller til akkurat rett tid og sted?
Stedsspesifikk (kontekstavhengig) informasjon, der celler har sin posisjon spesifisert i forhold til et romlig koordinat-system sin referanse, vil være ett gjennomgangstema. (38)
+'Forutgående mønster' (prepattern) som danner en mal eller skjelett for organisering av en påfølgende struktur. (39)
Angående informasjonslagring for dannelse av ulike kroppssystemer, gjelder at: Uten pre-eksisterende informasjon, kan ingen kroppsdannelse finne sted.
Om man kan generalisere, så vil et forhåndsmønster av noe slag, kanskje utviklet fra en type enda tidligere posisjons-system, styre genutvikling, som i sin tur styrer celle-oppførsel, som i sin tur styrer kroppsdannelse (morfogenese).
Det finnes en informasjons-strøm hvorfra de første utviklingsstadier, til dets resultat i endelige kroppsformer, som må bli nøyaktig dirigert og koreografert.
For skjelett-utvikling:
Et utvendig styrt elektrisk signal, spiller en rolle i å identifisere den korrekte lokalisering til fremtidige lemmer (Se kap 7A -her)
Spesifikke gener og deres produkter er aktive ved å produsere kjemiske signaler, integrert i et fire-dimensjonalt mønster-system. (42) Disse kjemiske signalene hjelper i å spesifisere utvikling av lemmene.
Disse signalene blir produsert i rett tid og sted (43) som indikerer et underliggende forhåndsmønster, som selv styrer hvordan genene uttrykkes.

Sentralnerve systemet
Hjernen og sentralnerve systemet oppstår fra Nevrale Progenitor Celler (NPC), som multipliseres og utvikles til nerveceller, og utvikler meget lange aksoner og dendritter som knyttes til målceller via synapser (48). Informasjonen, som styrer tilknytningen, anses å oppstå i NPC fra uttrykk via en kombinatorisk kode i proteiner (transkripsjonsfaktorer), som styrer andre geners uttrykksmåte. (49)
Denne koden må oppstå og bli nøyaktig styrt /kontrollert av pre-eksisterende informasjon (forhåndmønster). Resultat: "den intrikate koreografering" av celle-migrasjon, akson-styring, dendritt-vekst og synapsedannelse i sentralnerve-systemet. (50) Det må styres fordi det kreves at synapser forbindes 100% presist med hverandre. (51)
Spesifisering av informasjon
1. Forhåndsmønster: Forut for synapsedannelse mellom nabonevroner, inntreffer 'hotspotter' av spesialisering, som indikerer at forhåndsmønster forteller nevroner hvor synapser vil dannes. (52)
2. En transkripsjons-kode: Biokjemiske sendebud, utskilt langs de ulike kroppsaksene formodes å kommuniser posisjons-informasjon til NPCer. Det medvirker at de danner en kombinatorisk kode av transkripsjons-faktorer, som i sin tur forsyner informasjon som styrer nerveforbindelser. (53)
Celler som blir spesialiserte (f.eks. øyet) krever nøyaktig samordning av komplekse regulerende mekanismer (J-P Saint-Jeannet, prof. Toulouse universitet).
 3. En histonkode: kromosomer er sammensatt av strenger av DNA som er rullet rund histon-proteiner. Endringer i disse histonene inngår i aktivering/deaktivering av gen-aktivitet. Modifikasjoner av histon-proteiner forsyner en histon-kode, som multipliserer det kombinatoriske potensialet og således den felles informasjonskapasiteten til genene.
3. En histonkode: kromosomer er sammensatt av strenger av DNA som er rullet rund histon-proteiner. Endringer i disse histonene inngår i aktivering/deaktivering av gen-aktivitet. Modifikasjoner av histon-proteiner forsyner en histon-kode, som multipliserer det kombinatoriske potensialet og således den felles informasjonskapasiteten til genene.
4. En celle-overflate kode: Karbo-hydrat proteiner komplekser i celle overflaten har et stort informasjons-potensial. Endringer i deres uttrykk korrelerer med hendinger i løpet av utvikling av nervesystemet (56) og hindring ødelegger denne utviklingen. (57) (Se kap 7B -her)
5. En bioelektrisk kode som består av elektriske signaler i visse områder i det utviklende embryoet, bl.a. forutsier det påfølgende mønster i nervesystemet (58) og å gripe inn i dette fører til defekter i utviklingen.(59)
DNA og ulike epigenetiske kilder utenom DNA forsyner altså kodet informasjon som både er sofistikert og integrert sammen i sine aktiviteter og det hjelper til sammen å styre dannelsen av nervesystemet.
Utvikling av hjerte og kretsløps-system
Utvikling av hjerte og kretsløps er en forunderlig og presis sammensatt serie av molekylære og morfogenetiske begivenheter" (hjertespesialist D. Srivastava -62)
Hjertets utvikling oppstår fra en region i hjerte-dannende celler i det tidlige embryoet, kjent som det første hjerte-feltet 864) Disse cellene utvikler, migrerer og dannes til en lineær tube.
Senere migrerer en gruppe celler (det andre hjerte-feltet) til hjerte-tuben og tillater den å bli forlenget, og så loope til høyre, utvide seg og bli omformet til hjertekamrene (atrier og ventrikler) (64)
Celler fra den nevrale tuben migrerer også til hjerte-tuben og kreves for dannelse av hjerte-klaffene, og avdelinger som skiller oksygenrikt fra oksygenfattig blod. (65)

Et meget stort antall gener er involvert i disse prosessene, med et nettverk av transkripsjons-stier, som aktiverer hverandres uttrykk. (67) Kathiriya et al. beskriver hvordan kardio-transkripsjonsfaktorer koreograferer' uttrykket til tusener av gener ved hvert stadium av hjerte-utviklingen, ved å interagere med kofaktorer og binde til en konstellasjon av regulerende DNA-elementer. Disse nettverkene samvirker for å 'dirigere' den sekvensielle iverksetting av kardio-genuttrykks programmet. (68)
Arterie og ventrikkel-kamrene trekker seg sammen sekvensielt, som er vitalt for effektiv for utpumpingen av blod(69). Det utføres ved mikroskopiske muskel-proteinceller, som presist er registrert sammen. Elektriske impulser fra pacemaker-celler sprer seg raskt gjennom arterien , og resulterer i samtidig sammentrekning av den.
Arterier trekker seg sammen før hjertekamrene, så spres elektrisk strøm gjennom hjertekamrene, gjennom et spesialisert hurtig åre-system, som fører til sekvensiell sammentrekning av hjertemuskelen.
For at det kardio-vaskulære systemet skal fungere skikkelig, så er det ikke en uniform morfologi hos arterier, vener og lymfeårer, men de har distinkt ulike strukturelle trekk, for å utføre ulike oppgaver. (78)
Arterier har f.eks. tykke indre vegger for å holde stand mot blod som presses ut fra hjertet i høyt trykk. Vener har tynne vegger, og er konstruert for å la hvite blodlegemer passer gjennom seg. De har også ventiler for å hindre tilbakestrømning av blodet til kapillærårene. (79)
Arterie og vene-spesifiseringen krever at 'korrekt romlig og temporært uttrykk av mange gener er samordnet av en relativt stort sett av transkripsjonsfaktorer,' (80) som 'jobber samordnet sammen' (81)
 Informasjonens natur
Informasjonens natur
Om organismer ikke er intelligent designet, så må de være satt sammen av ikke-intelligente mekanismer. (88) Mekanismer som flikker/retter på ting er i sentrum av moderne evolusjonsteori.Men ovenfor så vi mange eks. på intrikat samordning for å drifte de samtidig koordinering av ulike organer og organsystemer, integrert med reproduktiv oppførsel.
Det er ikke bare en genetisk kode, men mange koder sammen som gir rik kombinasjons-kapasitet for informasjonslagring. I tillegg til ovennevnte liste med koder, kunne også nevnes spleisekodene, cytoskjelett-koder, apoptosis (programmert celledød) kode og ubiquitin-(beskyttelses) koden (89). Det er videre kjent at biologiske koder ikke bare overfører informasjon, men også fortolker den. Samt at den biologiske koden omfatter hvordan en kan produsere nye sendere og mottakere. (90) De anerkjenner også programmeringsaspektet, at den er skrevet på et språk som gjenstår å bli avkodet. Og det er ikke bare lineære sekvenser av instruksjoner, men et program med subrutiner, med prosedyre-kall, looper og all kompleksitet som det innebærer. (91) Således er premisset om enkel lineær informasjonsflyt i ferd med å skygge banen for en ny utviklingsmodell, som involverer biologiske nettverk innen hierarkiske lag for rutine-kall. Informasjon flyter i flerfoldige retninger både innen og mellom lagene. (92)
Fysikeren og ingeniøren W. Gitt hevder at det ikke er vist eksperimentelt at informasjon kan oppstå spontant i materie. (94) Det er bare gjennom styring avt prosesser "går motsatt i forhold til naturloverenes retninger, for å oppnå mål som ikke-styrte prosesser aldri kan oppnå. Mer om det her.
 Delingsmønstre
Delingsmønstre
Det romlige arrangementet av celler innen embryoet i det de deler seg, er kjent som delingsmønsteret. Ulike dyregrupper har bemerkelsesverdige fundamentale forskjeller i disse mønstrene, som flg. eks. demonstrerer:
a. Ctenophora (Kamskjell 113)
De første to delingsmønstre er i det vertikale (meridional) planet. Den tredje nesten-vertikale deling resulterer i en kurvet plate med 8 større celler (makromerer). Så følger en horisontal deling som gir opphav til små celler (mikromerer) på den konkave siden av makromerene. Mikromerene deler seg flere ganger og makromerene produserer et nytt sett av mikromerer.
 b. Bløtdyr (snegler 114)
b. Bløtdyr (snegler 114)
Bløtdyregg fremviser en spiral-kløvning, der hvert fjerdel av mikromerer roteres til høyre/venstre for makromerene i alternative delinger. De første to delinger er designet på samme vis som for Ctenophora (kamskjell). I følgende deling deler hver av cellene (makromerer) seg for å danne en mikromer. denne prosessen videreføres for videre å danne kvartaler av mikromerer, som produserer ytterligere delinger (Fir9.7)
Fig. 9.7 og 9.8 Bløtdyr og insekt
c. Arthropoda: insekter (116)
I bananfluen (drosophila), deler kjernen seg, men forblir i den samme felles cytoplasmaen (syncytium) (Fig. 9.8) (117) Gradvis folder egg-membranen seg innover for å avdele hver celle kjerne i enkeltceller. Dette mønster med tidlig syncytikal utvikling er felles for de fleste insekter.
 d. Virveldyr
d. Virveldyr
Delingsmønsteret atskiller seg signifikant mellom grupper av fisk, frosk, kylling og mus -som alle har distinkte mønstre for celledeling, som alle igjen er ulike de ovenfor nevnte delingsmønstre.
Den avgjørende betydning av tidlig utvikling blir avslørt gjennom eksperimenter der individuelle celler fra tidlige oppdelingsmønstre blir ødelagt. I bløtdyr embryoer fører dette til tap av voksenstrukturer som stammet fra ødelagte celler.
Evolusjonister har en tendens til å nedprioritere eller overse betydningen av disse tidlige stadier, men det skulle understrekes at i slike dyr, er de tidlige stadier grunnleggende for den påfølgende utvikling av former. I noen andre dyregrupper blir cellens mål/skjebne avgjort senere.
Hvordan kan fundamentalt ulike delingsmønstre spores til en felles stamfar?
Den foreslåtte eksistensen av en slik synes å trykkes ned over bevisene, heller enn å stamme fra dem!
Fordelingen av genprodukter viser fundamentale diskontinuiteter mellom basisformene. Fig 9.9 (119)
Fig. 9.9 Zebrafisk, amfibium og sjøpinnsvin
Hox-gener og det fylogenetiske stadiet
Det har vært stilt forslag ved at nye kroppsformer kunne dannes ved at hox-gen klustre skulle kunne bidra til nye kroppsformer, men det er atskillige problemer og hull i disse argumentene:
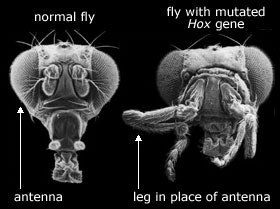 i) I tallrike dyrerekker (eks. bløtdyr) kan denne formodede utviklings-plastisitet ved artsbestemmelses-stadiet blir begrenset av grunnleggende tidligere begivenheter i embryoet.
i) I tallrike dyrerekker (eks. bløtdyr) kan denne formodede utviklings-plastisitet ved artsbestemmelses-stadiet blir begrenset av grunnleggende tidligere begivenheter i embryoet.
ii) Stadier (som det foreslåtte gastrula) kunne, som et nødvendig utviklingsstadium for disse dyrerekkene, forklares ved et felles design-mønster.
iii) Embryologen M. Richardson viste at det ikke er noen felles typisk stadium for dyre-ordener, slik at en kan utvide det som gjelder for virveldyr til virvelløse dyr. Her er det helt ulike utviklingsmekanismer mellom ulike dyrerekker, når de er mest like ytre sett. (125)
iv) Feilaktige diagrammer av Hox-klustre leder til misforståelser, i følge Denis Duboule, prof. i utviklings genetikk og genomikk ved både Ecole Polytechique og Geneve universitet. (126)
v) En slik ekskludering av intergenetiske sekvenser representerer en feilaktig oppfatning av at de ikke er viktige. (127)
vi) Duboule gjør en 'overraskende og pinlig' oppdagelse og konklusjon at 'virveldyr fremviser den mest tett organiserte Hox-gen klustre, mens de fleste andre dyr har et løst organisert eller splittet kluster.. (128)
Når arrangementet av Hox-klustre gjennomgås i detalj, så er det ingen overgangs-serie mot virveldyr mønsteret, noe som feiler å vise evolusjonær link til en felles stamfar.
vii) Igjen er det enda ukjent m.h.t. hvilken rolle Hox-ganet har i faktisk å generere fenotypen fra genotypen. Og dermed blir det som sagt spekulativt hvordan Hox og andre gen kan danne evolusjonært nye kroppsplaner.
Evolusjonistene R. Raff og Th. Kaufman innrømmer mangel på bevis for evolusjonære endringer i kroppsplaner, og sier at de fleste observasjoner ikke er gjort på organismer men på fossiler, for bare ved å gå tilbake til fossilregisteret får vi et syn for organismers faktiske evolusjonære historier.' (12) Om fossiler -se her.
Fossil registeret
 Raff og Kauffman gjør noen overraskende observasjoner:
Raff og Kauffman gjør noen overraskende observasjoner:
i) "Høyst komplekse dyr - pigghuder (sjøstjerner), trilobitter, leddyr og ulike klasser av bløtdyr dukker alle opp i Kambrium i betydelig variasjon og uten gjenkjente forfedre. (131)
ii) Overgangsformer er i hovedsak hypotetiske -selv de tidligste klasser av havdyr, er temmelig forskjellige fra hverandre. Det gjelder ikke bare pigghuder, hullene er generell og gjennomgående i fossilregisteret.
iii) Nye morfologiske strukturer dukker opp (amfibielemmer, vinger) og når de først dukker opp, er de fullt formet og ikke overgangsformer. (134)
iv) Hvordan ulike multicellulære kroppsplaner oppsto og skilte seg fra en felles stamfar, "Har vist seg å være fruktbar grunn for spekulasjoner, fordi det er så få fakta til å begrense innbilninger." (135)
Forsøk på å vise at de første forekomstene var mindre plutselige, har blitt motbevist av nye funn som gjenoppretter tidsskalaen. (136) Forsøk på å vise at rette geokjemiske forhold trigget kambrisk eksplosjon, er også lite overbevisende. Det er fordi de bare forsyner tillatende forhold, ikke årsaken til kompleks, spesifisert informasjon som er legemliggjort i den kambriske eksplosjonen.
F.eks. for Brontosaurus og Apatosaurus, rager bakre utkragning over bekken og baklemmer og høyeste ryggvirvel finnes der, det er for å avlaste maksimum belastning som finnes i den regionen, og ryggvinkelen avtar perfekt i forhold til kreftene som virker på den. Uten dette ville bena ryke under det enorme presset som skjelettet ble utsatt for. Dinosaur-skjeletter viser også evne til å endre form i forhold til dynamiske bevegelse.(141) Det kan observeres i store fossile gravplasser, men de er ikke gravplasser for feilende intelligent design.
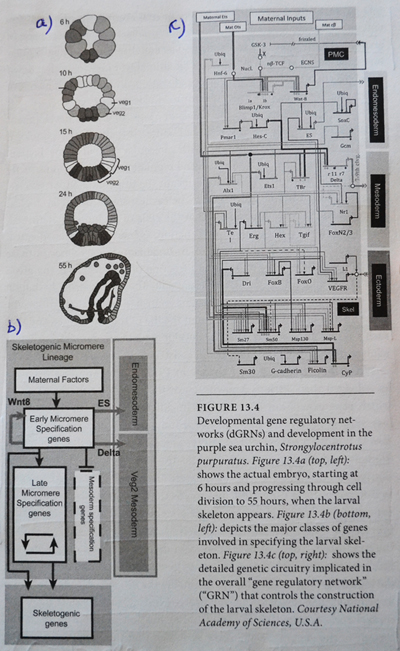 Taksonomisk sammenlikning av gen-utrykk
Taksonomisk sammenlikning av gen-utrykk
Det er bevis for særegenheter i ulike gen-uttrykks mønstre mellom taksonomiske grupper. F.eks. skiller mønster for benutvikling seg mellom insekter og edderkopper. (146) Mange transkripsjonsfaktorer som virker i amfibiets øye-utvikling, gjelder ikke for pattedyr, sammen med andre betydelige forskjeller mellom virveldyr. (147)
Det er altså bevis for eksistensen av ulike typer som er typer i naturen. Distinkte former inntreffer på ulike hierarkiske nivåer, og det er gjenkjennbart forskjell mellom de nærmeste former ved klare gap eller diskontinuiteter.
Spørsmålet for teistiske evolusjonister blir hvorvidt de vil basere seg på spekulasjonene til naturalismen, eller følge bevisene for design og diskontinuiteter dit de leder.
Referanser:
7. M. Lewin, "Morphogenetic Fields in Embryogenesis, Regeration and Cancer: Non-loal control of Complex Patterning," Biosystems 109, no.3 (2012), 243
37. J. Buchan, "Mr. Standfast," in The Complex Richard Hannay (Kondon: Penguin: 1992)
38. J. Jaeger et al, " On the Dynamic Nature on Positional Information," BioEssay
39. M. Jerka-Dziadosz et al, "Genetic Approaches to Ciliae Pattern Formation: Form Self Assembly to Morphogenesis," Trends in Genetics 6 (1990):41-45
42. Jean-Deniz Benazet et al, "Vertebrate Limb Development Moving from Classical Morphogen Gradients to an Integratede 4-Dimensional Patterning System," Cold Spring Harbor Perspectives in Biology 1, no. 4(2009): a001339
49. Fr. Polleux et al, " Transcriptional Regulation on Vertebrate Axon Guidance and Synapse Formation," Nature Reviews Neuroscience 8, no.5 (2007):331-340
50. D.A. Colon-Ramos, "Synapse Formation in Developing Neural Circuits," Current Topics in Developmental Biology 87 (2009):53-79
51. R. Jüttner et al, "Molecular Analysis of Axional Target Specificity target Specificity and Synnapse Formation," Cellular and Molecular Life Sciences 62, no.23 (2005):2811-2827
52. A.M. Craig et al, "Synnapse Formation and Maturation," in Synapses red. W.M. Cowan et al (Baltimore: John Hopkins University Press, 2001), 571-612
53. Polleux et al, "Transcriptional Regulation," 331-340
57. R. Bourrillon et al, " Cell Surface GlucoProteins in Embryonic Development," Yale Journal of Biology and Medicine 9, no. 6 (1937): 540
64. D. Franco et al, "Contemporary Cardiogenesis: New Insights into Heart Development," Cardiovascular Research 91, no. 2(2011): 183-184
65. Miquerol et al, " Establishment of the Mouse Ventricular Conduction System."
67. E. N. Olson, "Gene Regulatory Networks in the Evolution and Development of the Heart," Science 313, no. 5795 (2006): 1922-1927
68. I.Y Kathiriya et al, "Investigating the Transcriptional Control of Cardiovascular Development," Circulation Research 116, no. 4 (2015): 700
69. Miquerol et al, "Establishment of the the Mouse Ventricular Conduction System. "
78. M. Corado et al, " Signalling Pathways in the Specification of Arteries and Veins," Vascular Biology no. 11, (2014), 2372-2377
79. J. E. Fish et al, " The Molecular Regulation of Arterievenous Specification and Maintenance," Dev. Dynamics 244, no. 3(2015) 391-409
80. Corada et al, "Signalling Pathways," 2373
81. Fish et al, "Molecular Regulation of Arteriovenous Specification," 391
88. Fr. Jacob, "Evolution and Tinkering," Science 196 (1977): 1161-1166; A. Meier et al, "Evolution and Tinkering Gene Regulatory Landscapes,_" Current Biology 25, no. 7 (2015): R285-R288
89. M. Barbieri, " A new Theory of Development: The Generation of Complexity in Ontogenesis," Philosophical Transactions of the Royal Society A 374, no. 2063 (2016: 20150148
90. J.H.E. Cartwright et al, "DNA as Information At the Crossroads between Biology, Mathematics, Physics and Chemistry," Philosophical Transactions of the Royal Society A 374, no. 2063 (2016: 2015.0071, 7
91. Cartwright et al, "DNA as Information", 3
92. S. Franklin et al, "Genomes, Proteomes, and the Central Dogma," Circulation: Cardiovascular Genetics 4, no. 5 (2011): 576
113. C. Sardet et al, "Reproduction and Development in Ctenophores," i Experimental Embryology ind Aquatic Plants and Animals, red. H.J. Marthy (New York: Plenum 1990)
114. N.H. Verdonk et al, "Early Development and the Formation of the Germ Layers," i The Mollusca: Vol. 3: Developmen, red. N.H. Verdonk et al. (New York: AcademicPress , New York, 1983), 91-122
116. J. Kawana et al, "Insecta," i Invertebrate Embryology, red. M. Kume et al (Belgrade, Yogoslavia: NOLIT, 1968), 405-484
117. etter Kawana og Takami, "Insecta."
119. R.C. Range et al, "Integration of Canonical and NonCanonical Wnt Signalling Pathways Patterns .. PLoS Biology 11, no. 1(2013): e1001467
125. M.K. Richardson et al. "THere is no Highly Conserved Embryonic Stage in the Vertebrates: Implication for Current Theories of Evolutional and Development," Anatomy and Embryology 196, no. 2 (1997): 91-106.
127. D. Duboule, "The Rise and Fall of Hox Gene Clusters," Dev. 134, no.14 (2007): 2549
128. D. Duboule, "The Rise and Fall of Hox Gene Clusters," Dev. 134, no.14 (2007): 551
129. R.A. Raff et al, Embryos, Genes and Evolution (London: MacMillian, 1983), 25
131. Raff og Kaufmann, Embryos, Genes and Evolution, 31
134. Raff og Kaufmann, Embryos, Genes and Evolution, 54
135. Raff og Kaufmann, Embryos, Genes and Evolution, 30
136. S. Zamiora, "Middle Cambrian Echinoderms from North Spain Show Echinoders Diversified Earlier in Gondwana," Geology 38, no. 6 (2010), 507-510
141. eesc.columbia.edu/courses/v1001/twomed.html
147. Thomas Floss et al, "Pleiotropic Effects in Eya3 Knockout Mice," MBC Developmental Biology 8, no. 1 (2008):1.
Oversettelse og bilder ved Asbjørn E. Lund